Published in: Proc. Natl. Acad. Sci. USA, vol. 97, no. 18,
pp. 9919-9924, (August 29, 2000):
"A Trans-Acting RNA as a Control Switch in Escherichia
coli: DsrA Modulates Function by Forming Alternative
Structures".
Richard A. Lease* and Marlene Belfort
Molecular Genetics Program, Wadsworth Center, New York State Department of
Health, and School of Public Health, State University of New York at Albany,
P.O. Box 22002, Albany, NY 12201-2002
* To whom reprint requests should be sent at the present
address: Department of Biophysics, Jenkins Hall, Johns Hopkins University, 3400
North Charles Street, Baltimore, MD 21218-2685.
E-mail: ral@jhu.edu.
Abstract:
Introduction:
Figure 1
(Primer extension analysis to monitor RNA stability):
Materials and
Methods:
Results:
DsrA
Affects the Turnover of Its Target mRNAs:
Figure 2 (DsrA-RNA
interactions in the middle-stem loop):
Computational
Analysis Supports a Putative Second DsrA-hns Interaction:
Structure
of DsrA Inferred from Nuclease Footprinting:
Figure 3 (Nuclease
footprinting of DsrA):
Change
in DsrA Structure Induced by hns RNA in Vitro:
Phylogenetic
Analysis:
Figure 4 (Phylogenetic
structure analysis):
Figure 5 (Models of
regulation):
DsrA
Interactions with rpoS RNA:
Discussion:
DsrA
Structural Dynamics:
What
Governs the Dynamic Interaction of DsrA with Its targets?
Acknowledgements:
References:
Additional
References:
Other Sites:
Feedback:
ABSTRACT:
DsrA is an 87-nucleotide regulatory RNA of Escherichia coli that
acts in trans by RNA-RNA interactions with two different mRNAs,
hns and rpoS. DsrA has opposite effects on these transcriptional
regulators. H-NS levels decrease, whereas RpoS (ss) levels increase. Here we show that DsrA
enhances hns mRNA turnover yet stabilizes rpoS mRNA, either
directly or via effects on translation. Computational and RNA footprinting
approaches led to a refined structure for DsrA, and a model in which DsrA
interacts with the hns mRNA start and stop codon regions to form a
coaxial stack. Analogous bipartite interactions exist in eukaryotes, albeit with
different regulatory consequences. In contrast, DsrA base pairs in discrete
fashion with the rpoS RNA translational operator.Thus, different
structural configurations for DsrA lead to opposite regulatory consequences for
target RNAs.
INTRODUCTION:
RNA plays a variety of regulatory roles in the cell, in addition to its
central function in translation processes. These activities are collectively
termed riboregulation (1). RNA-RNA interactions
provide a basis for sequence-specific RNA regulation of other RNAs in both
prokaryotes and eukaryotes (2, 3). DsrA, an 87-nt
untranslated RNA in Escherichia coli, is one such regulatory RNA. DsrA contains
regions of sequence complementarity to at least five different genes,
hns, argR, ilvH, rpoS, and rbsD (4), and it has been
demonstrated to regulate two of these genes, hns and rpoS, by
RNA-RNA interactions (4, 5).
H-NS is an abundant nucleoid-structuring protein with global
transcriptional repressor functions (6, 7). DsrA antagonizes H-NS
function by decreasing the levels of H-NS protein in the cell (4). In contrast,
the translation of RpoS, the stationary phase and stress-response
sigma factor (8), is
enhanced byDsrA, especially at low temperatures (9). Thus, DsrA has
opposite effects on these two targets, both mediated by RNA-RNA
interactions, with global regulatory consequences for the
transcriptional state of the cell. Whereas the mechanism of DsrA
action at hns is not known, DsrA binds the translational
operator of rpoS (4, 5) to open a stable
stem-loop of rpoS RNA (10), enabling access
to the Shine-Dalgarno sequence and thus enhancing translation.
Structure predictions based on thermodynamic criteria suggest that DsrA
consists of three stem-loops, the third of which is the transcription
terminator of DsrA (Fig.
1 A; ref. 11).
Figure 1:
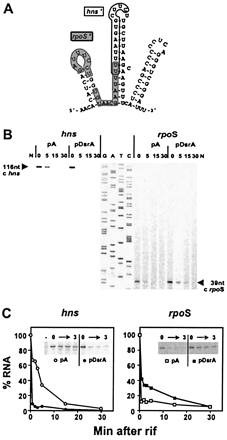
Fig. 1. Primer extension analysis to monitor
RNA stability. (A) Computer-predicted structure of DsrA RNA (11). The
rpoS-complementary region (rpoS') is shown on a gray background;
the hns-complementary region (hns') is outlined (4). (B) Primer extension
analysis of hns and rpoS mRNAs. RNA from cells containing a
plasmid (pDsrA) was compared with a vector control (pA). Numbers indicate the
time (min) after rifamycin addition (0 min). N, no-RNA control. A DNA sequencing
ladder (G, A, T, C) was used as a mobility marker. (C) RNA stability plots. cDNA
levels from primer extension analyses were quantified by PhosphorImager and
plotted relative to the zero time point for each experiment. The decrease in
hns mRNA half-life in the presence of excess DsrA is >8-fold. The
increase in rpoS mRNA stability in the presence of excess DsrA is
~3-fold. Gels inset into the graphs are primer extension analyses of 0-3 min
time points.
The hns complementary region, in the center of the
molecule, resides within the predicted second stem-loop, whereas the
rpoS complementary region occupies the predicted first
stem-loop and the base of the second stem (4, 5). However, no additional
structural data are currently available for DsrA. Mapping of the
different regions of complementarity on the structure of DsrA is
important for understanding how such a small RNA molecule interacts
with multiple targets, and how such interactions might be
regulated.
Here we show that the basis of DsrA regulation at hns involves
enhanced turnover of hns mRNA, perhaps by blocking H-NS translation,
whereas the translational stimulation of RpoS involves stabilization of
rpoS mRNA. The secondary structure of DsrA, determined in theabsence and
presence of hns RNA in vitro, suggests a distinct DsrA-rpoS
interaction involving stem-loop 1, whereas a two-part DsrA-hns
interaction involves stem-loop 2. The DsrA-hns RNA interactions
are proposed to circularize hns mRNA and define a structural
basis of DsrA activity at hns. Interestingly, similar
structures can be drawn for DsrA:argR and DsrA:ilvIH
RNA-RNA interactions. Taken together, these data show DsrA to be a
dynamic RNA molecule that changes its structure and interactions to
play different regulatory roles.
MATERIALS AND METHODS:
Strains, Plasmids, and Culture Conditions:
Computational Analyses:
RNA Stability Primer Extension Assay:
DsrA Substrate Construction:
DsrA and hns RNA Preparation In
Vitro:
Labeling of Purified DsrA:
Nuclease Cleavage Assays:
RESULTS:
DsrA Affects
the Turnover of Its Target mRNAs:
Previously, DsrA was shown to decrease the levels of H-NS protein, but not
steady-state hns mRNA levels (4). Because H-NS
represses its own transcription (6), a decrease in H-NS
protein should lead to increased hns mRNA levels. How, then, does
DsrA block synthesis of H-NS? Curiously, DsrA decreases the
steady-state mRNA levels of a Dhns
null mutant, in which autoregulation is abolished (4). Therefore, we
hypothesized that DsrA exerts an effect on hns RNA turnover.
To test the effects of DsrA on RNA turnover, the initiation of RNA
synthesis was inhibited with rifamycin and RNA levels were assayed by
primer extension analysis. When overproduced from a plasmid (pDsrA),
DsrA substantially decreased hns mRNA stability relative to
vector (pA) controls (Fig. 1 B
Left). Quantitation of mRNA levels taken from 1 to 30 min
after addition of rifamycin shows that hns mRNA levels dropped
immediately after rifamycin addition, with a sharp decrease in mRNA
half-life, from ~4 min to <0.5 min (Fig. 1 C Left), a
greater than 8-fold difference.
DsrA acts at rpoS by enhancing RpoS translation (9) via RNA-RNA
interactions that antagonize the rpoS translational operator
(4,
5, 10). Here we show that
DsrA overproduction alsoincreases rpoS mRNA stability (Fig. 1 B Right).
The levels of rpoSmRNA were stabilized relative to the control. Although
the relative stabilization was difficult to determine because of the short
half-life of rpoS mRNA, we estimate an approximately 3-fold increase
in rpoS stability (Fig. 1 C Right),
in sharp contrast to the effect of DsrA on hns mRNA (Fig. 1 C
Left).
Figure 2:
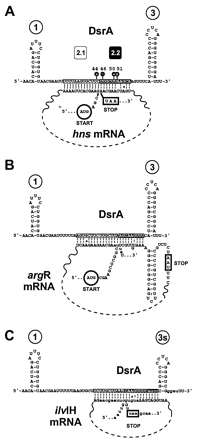
Fig. 2. DsrA-RNA interactions in the
middle-stem loop. (A) Computer sequence alignment of the hns coding
sequence depicted as a coaxial stack with regions of DsrA. The two regions of
DsrA complementary to hns are depicted as boxed (region 2.1) and as white
letters on a black background (region 2.2). Nuclease susceptibility (black
lollipops) and numbers over stem-loops are as in Fig. 3. (B and C) Other
RNAs modeled as coaxial stacks. The argR (B) and ilvH (C) RNAs of
E. coli with DsrA complementarity. Because of overlap in the DsrA complementary
sequences, there is potential for different pairings within coaxial stacks
(lowercase letters). Vertical lines represent base pairs verified by
compensatory mutagenesis (4); dashed vertical lines
represent base pairs predicted by informatic of biochemical analyses; a colon
(:) represents a G:U base pair; O represents a G:A base pair. Start codons are
circled; stop codons are boxed.
Computational
Analysis Supports a Putative Second DsrA-hns Interaction:
The DsrA-complementary region within hns described (Figs. 1 A and 2 A, boxed
sequence) was initially found by computer alignment and verified by
compensatory mutagenesis (4). A computational
reexamination of the hns coding sequence revealed a second potential
interaction of hns with DsrA (Fig. 2 A, white
letters on a black background). Interestingly, whereas the previously described
complementary sequence is near the start codon for hns translation, the
secondregion is near the hns stop codon (Fig. 2 A). Base
pairing between DsrA and the two regions of hns would form a
contiguous coaxial stack, looping out the middle part of hns
mRNA (Fig. 2).
The putative interaction between DsrA and the hns stop codon region
encouraged us to reexamine other genes by computer sequence alignment with DsrA.
As with hns, DsrA has the potential to pair with both
argR and ilvIH mRNAs near the start codon (ref. 4;Fig. 2
A-C, boxed sequences). Strikingly, computational
analysisrevealed that in analogy to hns, DsrA also contains extended
sequences with potential to base pair with both the argR and ilvI
stop codon regions (compare Fig. 2
A-C, white letters on a black background).Similarly to
hns mRNA, coaxial stacks can be modeled on the two-part
DsrA:argR and DsrA:ilvIH mRNA sequence complementarities (Fig. 2 B and C),
lending a common theme to all three potential RNA-RNA interactions involving the
DsrA middle stem-loop. In all three cases, the stop codons are near the putative
DsrA interactions. Although the effects of DsrA on argR and ilvlH
are not known, the theme of base pairing interactions with the start
and stop codon regions by the central region of DsrA could represent
a general mode by which DsrA regulates its target
genes.
Structure of
DsrA Inferred from Nuclease Footprinting:
DsrA interacts with the mRNAs of both hns and rpoS by
RNA-RNA interactions that involve distinct regions of DsrA (4, 5).To determine the
structural basis of these phenomena, the conformation of DsrA in solution was
deduced by RNase footprinting (23) with
single-strand-specific endonucleases, RNase T1 and S1
nuclease, and the double-strand-specific RNase V1. For
this purpose, we produced a DsrA substrate with precisely defined 5'
and 3' ends via a double ribozyme construct with DsrA in the middle
(15).
Cleavage by nucleases can induce conformational rearrangements
that lead to secondary cleavages, complicating RNA structural
analysis (23).
Accordingly, it was necessary to compare nuclease footprinting of
both 5'- and 3'-labeled substrates to obtain a consensus structure
(Fig. 3 A and
B).
Figure 3:
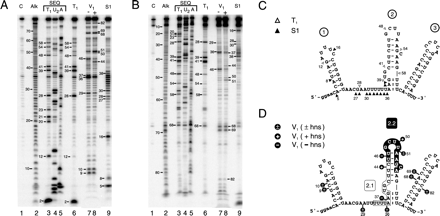
Fig. 3. Nuclease footprinting of DsrA. DsrA
substrate RNA produced and end-trimmed in vitro was labeled at the 5' or
3' end, then treated with ribonucleases. Digestion products are numbered by
phosphodiester bond position from the 5' end. (A) Footprinting of 5'-labeled
DsrA. (B) Footprinting of 3'-labeled DsrA. Lane numbers are given below, with
reactions specified above. Lane C, untreated control RNA; Alk, alkaline
hydrolysis ladder; lanes 3-5, sequencing reactions performed under denaturing
conditions; and lanes 6-9, specific nuclease footprinting reactions performed
under nondenaturing conditions. Lanes 3 and 6, RNase T1; lane 4,
RNase U2; and lane 5, RNase A; lanes 7 and 8, double-strand-specific
RNase V1 reactions performed in the absence or presence,
repectively, of unlabeled hns RNA; and lane 9, S1 nuclease. (C and D)
Predicted secondary structure of DsrA based on single-strand- and
double-strand-specific nuclease susceptibility. (C) Single-strand-specific
nuclease susceptibility. Open D, T1;
and solid D, S1 nuclease. (D) Double-strand-specific
nuclease susceptibility. The hns-complementary region characterized
(4) is shown boxed
(region 2.1) whereas the newly proposed hns-complementary sequence
(region 2.2) is shown in white letters on a black background. Cleavage sites are
indicated by black lollipops. +, cleavage only in the presnce of
hns RNA; -, cleavage only in the absence of hns RNA; +/- cleavage
occurs in the presence or absence of hns RNA; and :, partial
double-strand character.
RNase T1 cleaves after unpaired guanosine residues, whereas
residues within helices are unaffected (Fig. 3 A and
B, compare lanes 3 and 6; Fig. 3 C). For
example, nucleotide positions 39 and 41 are especially sensitive to
RNase T1 when DsrA is in eitherdenatured or native form, suggesting
that this region is singlestranded under native conditions. In contrast,
positions 20, 21,and 23 are cleaved only under denaturing conditions, and
musttherefore be double stranded in solution. The structural informationderived
from these analyses is summarized in Fig. 3 C (D).
RNase V1 cleaves double-stranded RNA of 4-6 bp, although not
all such sequences are cleaved (23, 24). Digestion with
RNase V1 confirms the existence of several regions of
double-stranded structure within DsrA, notably stem-loop 1 (Fig. 3 A lane 7,
nucleotides 9-10; Fig. 3
D) and stem-loop 3 (Fig. 3 A, B,
and D, nt 68, 69, and 82; additional data not shown),
confirming the computed thermodynamic predictions. Comparison of
T1 and V1 nuclease cleavage patterns suggests the
structure for stem-loop 2 depicted (Fig. 3 C,
positions 39 and 41, and Fig. 3 D,
positions 37 and 46), with an unstructured stretch between stem-loops
1 and 2, except at position 29 (compare Fig. 3 C and
D).
We further characterized DsrA secondary structure by treatment with S1
nuclease, which cleaves single-stranded RNA without sequence preference (23). The S1 nuclease data
confirmed the loop region for stem 1, because cleavage at position 16
(Fig. 3 A and
B, lanes 9) is consistent with RNase T1 and
V1 analyses and thermodynamic predictions (Fig. 3 C and
D). DsrA was also cleaved by S1 nuclease in the predicted
single-strand region between stems 1 and 2 (Fig. 3 A and
B, lanes 9, nucleotide positions 27-38), with the
exception of position 29. This corresponds precisely with
V1 nuclease cleavage at position 29, indicating double strandedness,
although the pairing partner at position 29 is not known.
There is an apparent contradiction between the S1 nuclease and RNase
V1 cleavage data for positions 36 and 37 (compare Fig. 3 C and
D). Positions 37-38 are therefore likely to be at least transiently
double stranded (Fig. 3
C and D, colons between residues). It is well known
that S1 nuclease is extremely active and can cleave regions of
double-stranded nucleic acid that transiently unpair or "breathe" (18, 25).
A 3'-labeled DsrA substrate behaves similarly to the 5'-labeled substrate,
verifying this analysis (Fig. 3 B). In summary,
the consensus secondary structure map (Fig. 3 C and D) confirms
previously predicted stem-loops 1 and 3 (11), whereas suggestinga
different structure for stem-loop 2. Furthermore, the data suggesta dynamic or
partial double-strand character at the base of stem-loop2.
Change
in DsrA Structure Induced by hns RNA in Vitro:
As mentioned, DsrA forms a RNA-RNA interaction with hns mRNA,
decreasing the levels of H-NS protein (4), evidently by
enhancingRNA turnover (Fig. 1). To examine the
structure of this DsrA-hns RNA complex, excess unlabeled
hns RNA was mixed with end-labeled DsrA and the complex was
treated with RNase V1 to look for variations in
double-stranded RNA regions. Whereas one main region of double-strandedRNA
exists in the central part of DsrA in the absence of hns RNA(Fig. 3 A, lane 7,
nt position 46), two enhanced nuclease-susceptible regions of
double-stranded RNA form on addition of hns RNA (Fig. 3 A and
B, compare lanes 7 and 8, nt positions 44 and 50-51). An equivalent
result was seen by using 3'-labeled substrate (Fig. 3 B, compare
lanes 7 and 8, nt positions 44 and 50-51; additionaldata not shown). The changes
in DsrA associated with the hns RNAinteraction are summarized (Fig. 2 A and 3 D, black
lollipops). As expected, the DsrA-hns RNA interaction at
position 44 of DsrA is within the previously described complementary
region (Fig. 3
D, region 2.1, boxed sequence; compare Fig. 1 A; (ref.
4). Interestingly,
the hns-induced double-stranded complex at positions 50-51
lies outside of this region, suggesting a second DsrA-hns RNA
interaction that includes the loop region of stem-loop 2 (Fig. 3D, nt
positions 50-51), in agreement with computer predictions (Fig. 2 A, and
Fig. 3 D,
region 2.2).
Phylogenetic Analysis:
DsrA sequences are available from three species, E. coli, S.
typhimurium, and K. pneumoniae. These sequences and their
predictedstructures (Fig. 4) are consistent
with our structural analyses.
Figure 4:
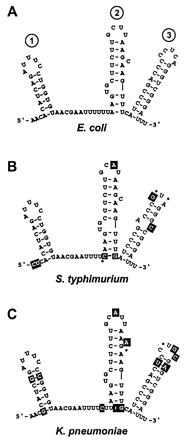
Fig. 4. Phylogenetic structure analysis.
Structure models of DsrA are given for E. coli (A), S.
typhimurium (B), and K. pneumoniae (C). Base changes relative to
the E. coli sequence are shown as white letters in a black box, and dots
indicate deletions.
Notably, sequence variations at the bottom of stem-loop 2 thatextend the
stem by one base pair are suggested in both the S.typhimurium and K.
pneumoniae sequences (Fig. 4 B and
C, white letters on a black background). Additional
changes in both S. typhimurium and K. pneumoniae
sequences relative to the inferred E. coli structure mostly
occur in regions predicted to be single stranded, or when in regions
predicted to be double stranded, are compensated by second-site
mutations that would restore base pairing (Fig. 4 B and
C, white letters on black background). Thisis the case for 23 base
changes of S. typhimurium and K. pneumoniaeDsrA relative to E.
coli DsrA. The only exception to these structural constraints on
sequence differences from E. coli DsrA is the single-nucleotide
substitution that creates a bulge in the third stem-loop of S.
typhimurium DsrA (Fig. 4B).
Figure 5:
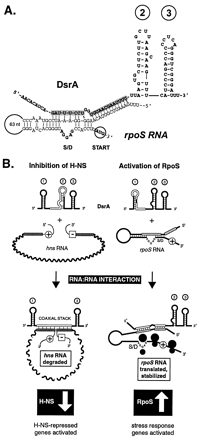
Fig. 5. Models of regulation. (A) DsrA shown
with the rpoS translational operator. The structure of the rpoS
translational operator (10) is shown with the new
DsrA structure model emphasizing the integrity of stem-loop 2. The
DsrA-rpoS RNA interaction also is described elsewhere (4, 5). (B) DsrA forms a
complex with hns mRNA (Left) and with rpoS mRNA (Right). The
hns- and rpoS-complementary regions in DsrA are outlined in white.
The start codons of hns and rpoS (plus sign in circle) and the
stop codon of hns (minus sign in box) are shown. The DsrA-hns RNA
interaction is shown below as a coaxial stack, with the second DsrA stem-loop
melted and the first and third stem-loops intact. In contrast, the DsrA complex
with rpoS mRNA has the first DsrA stem-loop melted outand the second and
third stem-loops intact. Stem-loops are numbered in circles.
DsrA Interactions with
rpoS RNA:
The interaction of E. coli DsrA with rpoS mRNA is also
consistent with the new DsrA structure model derived from RNA footprinting
analysis (Fig. 3 C
and D, and Fig. 5 A).
Here, in contrast both to the previous DsrA structure model (Fig. 1 A) and to
the DsrA-hnsmRNA interaction (Fig. 2 A),
stem-loop 2 is predicted to remain intact, whereas stem-loop 1 would
melt out and, together with the single-strand stretch between
stem-loops 1 and 2, contact rpoS mRNA (Fig. 3 C and
D, and Fig.
5). Thus, formation of RNA-RNAinteractions with DsrA targets would involve
distinct annealing processes and different DsrA conformations.
DISCUSSION:
DsrA RNA acts as an orchestrator of cellular gene expression by
simultaneously modulating the activities of at least two global
transcriptional regulators, H-NS and RpoS (Fig. 5 B), with
potential for direct interactions with the RNAs of at least three more genes,
argR, ilvIH, and rbsD (4). A revised secondary
structure model for DsrA, based on compelling RNA footprinting and
phylogenetic data (Figs. 3 and 4), accounts for its
differential regulationof hns and rpoS. Remarkably, DsrA is a
modulator of both RNA stability (Fig. 1) and translation
(4, 9). DsrA decreases H-NS
protein levels, whereas it increases RpoS levels, with a correspondingdecrease
in hns mRNA stability in vivo (>8-fold) and an increase
in stability of rpoS mRNA, approximately 3-fold (Fig. 1). As
discussed below, RNA stability may reflect the translational status
of the RNA.
DsrA Structural Dynamics:
DsrA performs disparate functions by forming alternative structures in
complex with target mRNAs (Fig. 2, 3, and 5). Whereas
footprinting confirmed stem-loops 1 and 3 of the structure originally
proposed for DsrA (11), the analysis predicts
a different structure for stem 2 (Fig. 3). Additionally, a
coaxial stack proposed between hns RNA and DsrA is postulated
as a basis for DsrA regulation of the hns message (Fig. 2). However,
whereas compensatory mutagenesis confirmed the 2.1 base-pairing
region (4), similar
experimentshave not proven the region 2.2 pairing (R.L., D. Smith, and
M.B.,unpublished results). Whereas this RNA-RNA interaction is clearlyseen in
vitro, the conditions are not yet understood for its formation in
vivo.
Interestingly, the DsrA-hns RNA interaction at the hns stop
codon (Fig. 2
A) is paralleled by potential RNA-RNA interactions at the
argR and ilvI stop codons, which would represent second
interactions for DsrA on these mRNAs (Fig. 2 B and
C). All three of these proposed RNA-RNA interactions at stop codons
contain a nonWatson-Crick G:A base pair (Fig. 2, open circles),
whichhas been seen in other RNA-RNA interactions (26), particularly in
coaxial stacks with single mismatches (27). The formation
of a coaxial stack is expected to stabilize both parts of these
RNA-RNA interactions (Fig. 2).
A coaxial contiguous helix between hns and DsrA would loop out the
central portion of hns mRNA (Fig. 2 A). In
this model, the first and third stem-loops of DsrA remain intact
whereas the central stem-loop melts during formation of the DsrA-mRNA
interaction. The effect of this RNA-RNA interaction at hns
would be to enhance the turnover of hns mRNA (Fig. 1 B and
C), perhaps by creating ribonuclease-sensitive sites in hns
mRNA, or by preventing translation, which would otherwise protect the mRNA from
degradation (Fig.5 B
Left). In this regard, however, we find that RNase III is
dispensible for DsrA activity at hns (R.L. and M.B.,
unpublished results). Cleavage at the 5' or 3' end could facilitate
degradation (28),
whereas cleavage at the 5' end could abolish translation by
eliminating initiation. Such a degradation model would prevent
translation and thereby offset increases in hns RNA levels
caused by reduced H-NS protein levels, because of H-NS autoregulation
(4). The proximity of
both hns and argR start and stop codons also suggest inhibition of
translation. Conversely, in eukaryotic systems, interactions between the 5' and
3' ends of mRNAs are proposed to enhance translation, although the requirement
for circularization in vivo has not been fully elucidated (29).
In contrast to the interaction with hns, it is the first stem-loop
of DsrA that is complementary to rpoS, and which forms the
RNA-RNA interaction with rpoS mRNA (Fig. 5 A, and Fig.
5 B Right; refs. 4, 5). Elucidation of the
structure of DsrA (Fig.
3) suggests that stem-loop 2 can remain intact while base paired
to rpoS mRNA (Fig. 5 A and
B). Although it is not yet known whether DsrA stabilizes
rpoS directly or indirectly, we favor the hypothesis that
enhanced translation reduces mRNA turnover (30).
What
Governs the Dynamic Interaction of DsrA with Its Targets?
Loops and single-strand stretches of RNA mediate initiation of RNA-RNA
interactions (31).
Sequences within DsrA complementary to both rpoS and hns reside in
regions of DsrA that appear either in loops or single-stranded regions (Figs. 3 C and
D, 4,
and 5 A),
suggesting that "kissing complexes" might form between DsrA and target mRNAs,
leading to the RNA-RNA interaction. The separation of two disparate functions of
DsrA (Fig. 1) into
distinct structural elements (Figs. 3 - 5) forms a basis both
for strand-strand interactions and potential regulation of these interactions
(Fig. 5
B).
RNA-binding proteins are also likely to be involved in the regulatory
activity of DsrA. Many small RNAs require a protein component for
activity, and function as ribonucleoprotein particles, as in the case
of tmRNA (32), RNase P,
and eukaryotic small nuclear ribonucleoproteins (33). The RNA chaperone Hfq
(HF-I)has been shown to be required for opening of the S. typhimurium
rpoS translational operator (10), and enhances DsrA action atrpoS RNA
(D. Sledjeski, personal communication). Hfq involvementhas also been proposed
for the activity of OxyS RNA at rpoS (34). Whereas
some hfq strains accumulate DsrA (D. Sledjeski, personal
communication), in our strains hfq-null mutations markedly
decrease levels of DsrA RNA, complicating analysis of the role of
this protein with DsrA in vivo (R.L. and M.B.,
unpublished results).
What advantage is there in having a RNA, not a protein, as a regulator?
Because of the specificity of base pairing interactions, DsrA can
target multiple independent sites with economy, because the small RNA
requires little energy to produce and is extremely stable, with a
half-life of > 30 min (R.L. and M.B., unpublished results).
Because of its small size and stability, DsrA could rapidly
accumulate in response to different environmental signals, to enhance
or inhibit turnover and/or translation of existing specific RNAs, and
persist to maintain this altered state.
The plethora of roles played by RNA, widely distributed and acting in
diverse ways, speaks not only to the flexibility of RNA as a
biopolymer but also to an evolutionary history rooted in a RNA or
RNA-protein world (33). Riboregulation is a form of regulation that, at minimum,
requires only RNA-RNA interactions. RNA governing turnover and/or translation of
RNA by changing both its conformation and that of its targets would be a form of
regulation that we could expect to see in such a RNA or ribonucleoprotein
world.
Acknowledgements:
We thank D. Smith and S. Lawrence for expert technical help, A.
Ferré-D'Amaré and R. Collins for providing plasmids and ribozyme suggestions, O.
Uhlenbeck for suggestions on 3' labeling,B. Cousineau and D. Sledjeski for
helpful discussions, B. Cousineau,S. Gottesman, S. Hanes, P. Masters, and S.
Woodson for suggestions on the manuscript, the Molecular Genetics Core facility
at WadsworthCenter for oligonucleotide synthesis and DNA sequencing, M. Carlfor
preparation of the manuscript, and J. Dansereau for help with figures. This work
was supported by National Institutes of HealthGrants GM39422 and GM44844 to
M.B.
References:
1. Eddy S, (1999) Curr. Opin. Genet. Dev. 9: 695-699.
2. Moss EG, Lee RC, and Ambros V, (1997) Cell 88:
637-646.
3. Wassarman KM, Zhang A, and Storz G, (1999) Trends
Microbiol. 7: 37-45.
4. Lease RA, Cusick ME, and Belfort M, (1998) Proc. Natl. Acad. Sci. USA
95: 12456-12461.
5. Majdalani M, Cunning C, Sledjeski D, Elliot T, and
Gottesman S, (1998) Proc. Natl. Acad. Sci. USA 95: 12462-12467.
6. Williams RM, and Rimsky S, (1997) FEMS Microbiol.
Lett. 156: 175-185.
7. Atlung T, and Ingmer H, (1997) Mol. Microbiol. 24:
7-17.
8. Hengge-Aronis R, (1996) Mol. Microbiol. 21:
887-893.
9. Sledjeski DD, Gupta A, and Gottesman S, (1996) EMBO J.
15: 3993-4000.
10. Brown L, and Elliott T, (1997) J. Bacteriol. 179:
656-662.
11. Sledjeski D, and Gottesman S, (1995) Proc. Natl.
Acad. Sci. USA 92: 2003-2007.
12. Zhang A, Rimsky S, Reaban ME, Buc H, and Belfort M,
(1996) EMBO J. 15: 1340-1349.
13. Chang ACY, and Cohen SN, (1978) J. Bacteriol. 134:
1141-1156.
14. Belfort M, Ehrenman K, and Chandry PS, (1990) Methods
Enzymol. 181: 521-539.
15. Ferre-D'Amare AR, and Doudna J, (1996) Nucleic Acids
Res. 24: 977-978.
16. McCoy MI, and Gumport RI, (1980) Biochemistry 19:
635-642.
17. Guo HCT, and Collins RA, (1995) EMBO J. 14:
368-376.
18. Sambrook J, Fritsch EF, and Maniatis T, (1989)
Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Lab. Press,
Plainview, NY).
19. Weber TR, (1985) Doctoral Thesis (Univ. of Illinois
at Urbana-Champaign).
20. England YE, and Uhlenbeck OC, (1978) Nature 275:
560-561.
21. Hinton DM, Baez JA, and Gumport RI, (1978)
Biochemistry 17: 5091-5097.
22. Kuchino Y, and Nishimura S, (1989) Methods Enzymol.
180: 154-163.
23. Moine H, Ehresmann B, Ehresmann C, and Romby P,
(1997) "RNA Structure and Function", eds. Simons RW, and Grunberg-Manago M,
(Cold Spring Harbor Lab. Press, Plainview, NY), pp. 77-115.
24. Ehresmann C, Baudin F, Mougel M, Romby P, Ebel J-P,
and Ehresmann B, (1987) Nucleic Acids Res. 15: 9109-9128.
25. Wiegand RC, Godson GN, and Radding CM, (1975) J.
Biol. Chem 250: 8848-8855.
26. Nagaswamy U, Voss N, Zhang Z, and Fox GE, (2000)
Nucleic Acids Res. 28: 375-376.
27. Burkhard ME, Turner DH, and Tinoco I, Jr., (1999)
in"The RNA World", eds. Gesteland RF, Cech TR, and Atkins JF, (Cold Spring
Harbor Lab. Press, Plainview, NY), pp. 233-264.
28. Arraiano CM, Cruz AA, and Kushner SR, (1997) J. Mol.
Biol. 268: 261-272.
29. Wells SE, Hillner PE, Vale RD, and Sachs AB, (1998)
Mol. Cell 2: 135-140.
30. Kushner SR, (1996) in "Escherichia coli and
Salmonella: Cellular and Molecular Biology", eds. Neidhardt FC, Curtiss R, III,
Ingraham JL, Lin ECC, Low KB, Magasanik B, Reznikoff WS, Riley M,
Schaechter M, and Umbarger HE, (Am. Sco. Microbiol., Washington, DC) pp.
849-860.
31. Zeiler BN, and Simons RW, (1998) in "RNA Structure
and Function", eds. Simons RW and Grunberg-Manago M, (Cold Spring Harbor Lab.
Press, Plainview, NY), pp. 437-464.
32. Karzai AW, Susskind MM, and Sauer RT, (1999) EMBO J.
18: 3793-3799.
33. Gesteland RF, Cech TR, and Atkins JF, (1999) "The RNA
World", (Cold Spring Harbor Lab. Press, Plainview NY), pp. 1-709.
34. Zhang A, Altuvia S, Tiwari A, Argaman L,
Hengge-Aronis R, and Storz G, (1998) EMBO J. 17: 6061-6068.
Additional References:
1. Altuvia, S., Wagner, E. G. H. "Switching On and Off with RNA".
Proc. Natl. Acad. Sci. U. S. A. 97: 9824-9826 (August 29, 2000).
2. "Nuclear
Polyanions as De-Repressors of Synthesis of Ribonucleic Acid".
3. "Mated Models of Gene
Regulation in Eukaryotes".
4. "Oncogenes as
Molecular Targets within Active Chromatin".
5. "The Role of Chromosomal
RNAs in Marking the X for Dosage Compensation".
Top of Page - Euchromatin Network - Current Research - Forums - Other Sites - Future Events -
For Further Information and Feedback:
E-mail: frenster@euchromatin.net
Phone: +1 650 367 6483
euchromatin: "the most active portion of the genome within the cell
nucleus".